- 品牌
- MCE,艾德莱,OriGene,abmart,ABW基质胶
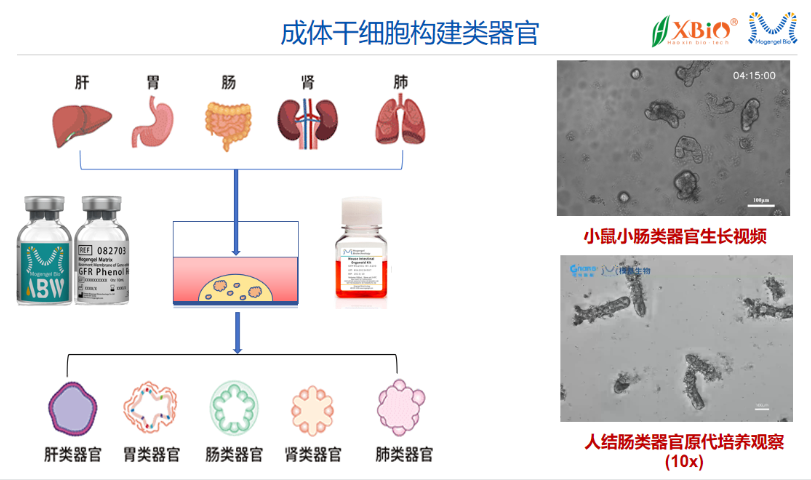
类***的培养为疾病模型的建立提供了新的思路。通过从患者的干细胞或组织中提取细胞,研究人员可以在基质胶中培养出与患者相似的类***。这些类***不仅能够模拟疾病的发生和发展过程,还能用于药物筛选和疗效评估。例如,在**研究中,类***可以用于评估不同化疗药物对肿瘤细胞的敏感性,从而为个性化***提供依据。此外,类***还可以用于研究遗传性疾病、***性疾病等,帮助科学家更好地理解疾病机制和寻找潜在的***靶点。尽管基质胶-类器官培养技术在生物医学研究中展现出巨大的潜力,但仍面临一些挑战。例如,如何提高类***的成熟度和功能性、如何实现大规模培养以满足临床需求等,都是当前研究的热点。此外,基质胶的来源和成分的复杂性也限制了其在临床应用中的推广。因此,未来的研究需要在优化培养基质、探索新型支撑材料以及提高类***的标准化和 reproducibility等方面进行深入探索。随着技术的不断进步,基质胶-类器官培养有望在再生医学、个性化***和药物开发等领域发挥更大的作用。基质胶中整合素配体的分布决定类器官的极性建立。西湖区免疫共培养基质胶-类器官培养性价比高
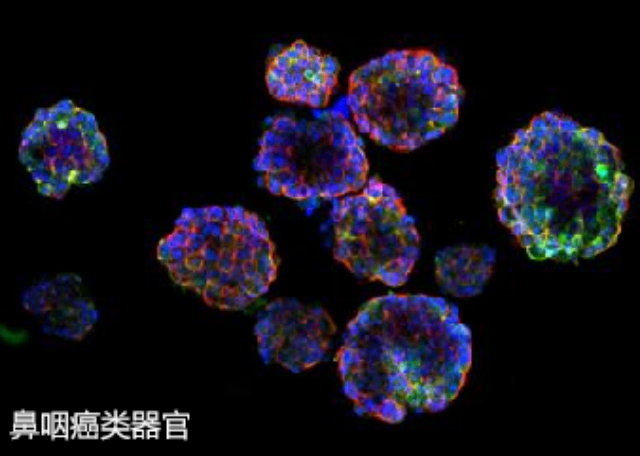
尽管类***培养技术在近年来取得了***进展,但仍面临一些技术挑战。首先,类***的标准化培养仍然是一个亟待解决的问题。不同实验室使用的培养基、基质胶浓度和培养条件可能存在差异,导致类***的形成和功能表现不一致。其次,类***的成熟度和功能性仍然有待提高。许多类***在培养过程中可能无法完全模拟真实***的复杂结构和功能,限制了其在疾病模型和药物筛选中的应用。此外,类***的长期培养和保存也是一个挑战,如何保持其活性和功能性是研究人员需要解决的问题。***,伦理问题也是类***研究中的一个重要考量,尤其是在使用人类干细胞时,如何确保研究的伦理合规性是必须重视的方面。桐庐低内毒素基质胶-类器官培养类器官与基质胶的互作机制尚需进一步深入研究。
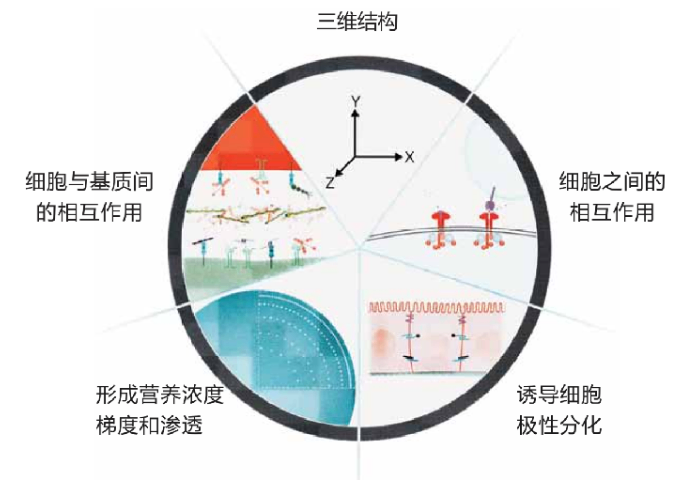
基质胶不仅是物理支架,更是重要的生长因子储库和调控系统。天然基质胶中含有多种内源性生长因子,包括bFGF、TGF-β、IGF等,这些因子在类***培养过程中发挥着关键的调控作用。更为重要的是,基质胶的三维网络结构能够实现对外源添加生长因子的可控释放。例如,通过将VEGF与基质胶中的肝素结合位点结合,可以***延长其半衰期并形成浓度梯度。在肠道类***培养中,这种缓释特性使得Wnt3a和R-spondin1等关键因子能够持续发挥作用,维持干细胞的自我更新能力。***研究还开发了多种生长因子递送策略,如微球包埋、亲和肽修饰等,进一步提高了生长因子在基质胶中的稳定性和生物利用度。这些进展为构建更加复杂的类***模型提供了重要技术支持。
尽管基质胶在类***培养中具有重要作用,但其来源和成分的复杂性也带来了一些挑战。例如,基质胶的批次间差异可能影响实验结果的 reproducibility。因此,研究人员正在探索基质胶的优化与改良方案,包括使用合成的细胞外基质材料或通过基因工程技术改造基质胶的成分。这些改良不仅可以提高类***的形成效率,还能增强其生物相容性和功能性。此外,研究者们还在探索如何通过调节基质胶的物理特性(如硬度、孔隙度等)来进一步优化类***的培养条件,以满足不同研究需求。类器官与基质胶的RNA测序需同步分析ECM相关基因。
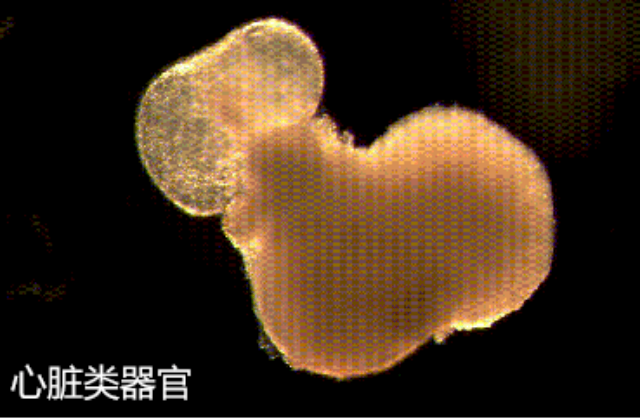
基质胶与生长因子的协同作用是类***培养成功的关键。基质胶不仅能物理性包埋生长因子,其某些成分(如肝素)还可通过结合和稳定生长因子来延长其活性。在肠道类***培养中,基质胶与Wnt3a、R-spondin1和Noggin的组合可维持干细胞特性;而在胰腺类***培养中,FGF10和EGF的添加时序对内分泌细胞的分化至关重要。***研究开发了生长因子梯度释放系统,通过将生长因子共价偶联到基质胶网络实现可控释放,显著提高了类***的成熟度和功能。通过显微操作可精确控制基质胶中类器官的初始接种位置。宁波基质胶-类器官培养批发厂家
类器官-基质胶模型可模拟肿瘤微环境中的免疫逃逸机制。西湖区免疫共培养基质胶-类器官培养性价比高
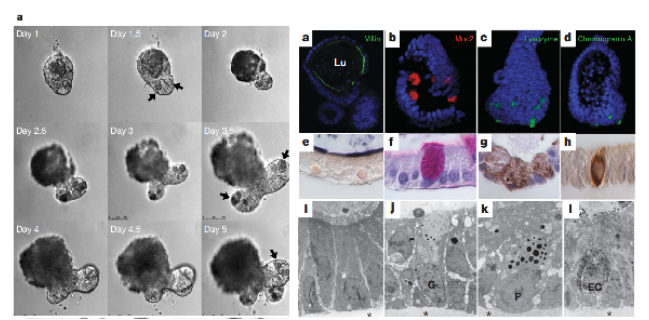
基质胶培养的类***为疾病研究提供了**性的模型系统。在**研究领域,患者来源类***(PDOs)保留原发**的组织结构和分子特征,已成为个性化医疗的重要工具。通过调节基质胶的硬度可以模拟不同阶段的**微环境,如较硬的基质(~8kPa)可诱导乳腺*的侵袭表型。在遗传性疾病研究中,囊性纤维化类***模型可以重现CFTR基因突变导致的病理变化。***进展是将基质胶类***与微流控系统结合,构建包含血管网络的复杂疾病模型,这为研究**转移和药物渗透提供了更真实的平台。此外,基质胶的组成调控还可以模拟特定病理条件下的ECM重塑,如肝纤维化中胶原沉积的增加。西湖区免疫共培养基质胶-类器官培养性价比高
- 浙江ABW基质胶-类器官培养实验步骤 2026-05-05
- 西湖区肠道基质胶-类器官培养价格怎么样 2026-05-05
- 萧山区干细胞分化基质胶-类器官培养怎么试用 2026-05-05
- 上城区ABW基质胶-类器官培养实验步骤 2026-05-05
- 肠道基质胶-类器官培养怎么试用 2026-05-05
- 湖州基质胶-类器官培养网上价格 2026-05-05
- 拱墅区细胞迁移与分化基质胶-类器官培养如何申请试用 2026-05-05
- 标准型金牌基质胶 2026-05-05